IMPACTS OF CLIMATE CHANGE
ON PLANT DIVERSITY IN SOUTH AFRICA
Methods Details for Succulent Karoo Bioclimatic Modelling
|
Projections of Spatial Shifts for the Dominant Biomes of South Africa by about 2050
Click each map to enlarge for details |
Maps A = Current Climate B = HadCM2 (Excluding sulphates) C = CSM (Excluding sulphates) D = HadCM2 (Including sulphates) |
Succulent Karoo Biome
The bioclimatic envelope of the Succulent Karoo has been modeled at present CO2 levels (365ppm), 475ppm CO2 equivalent and almost double pre-industrial levels (550ppm) using climate projections of three different general circulation models and these projections can be viewed by clicking the maps or links below.
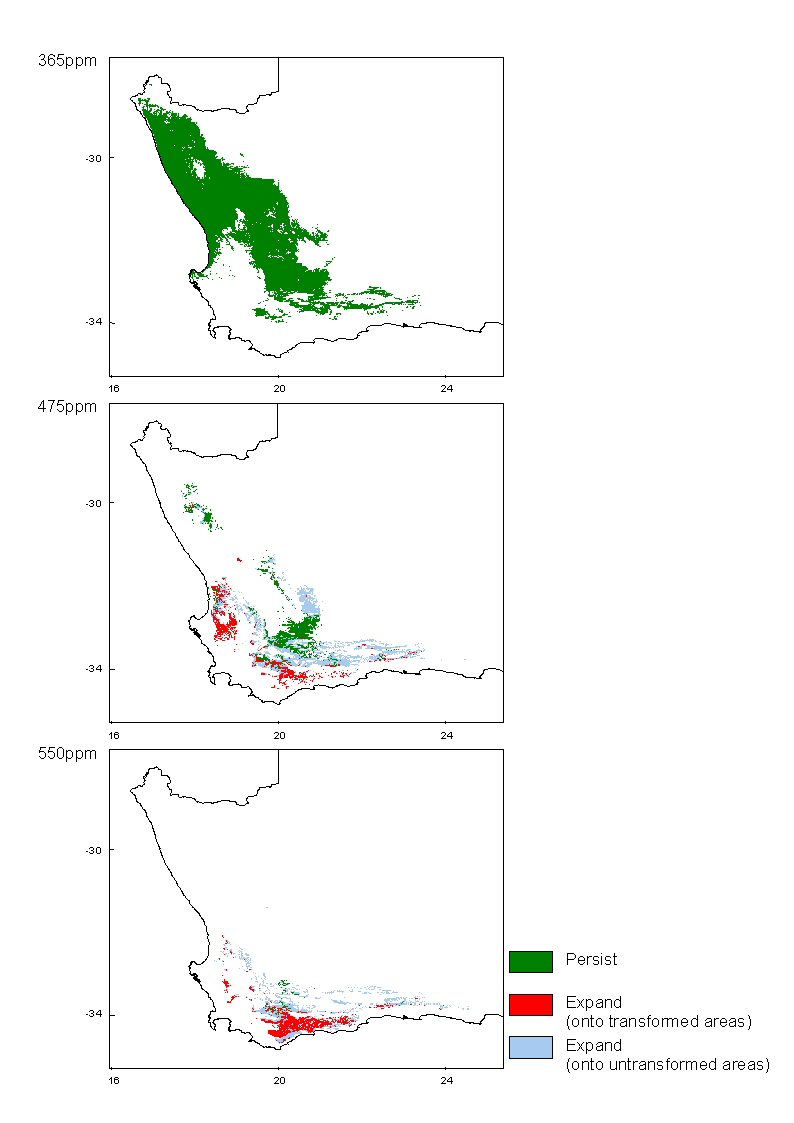
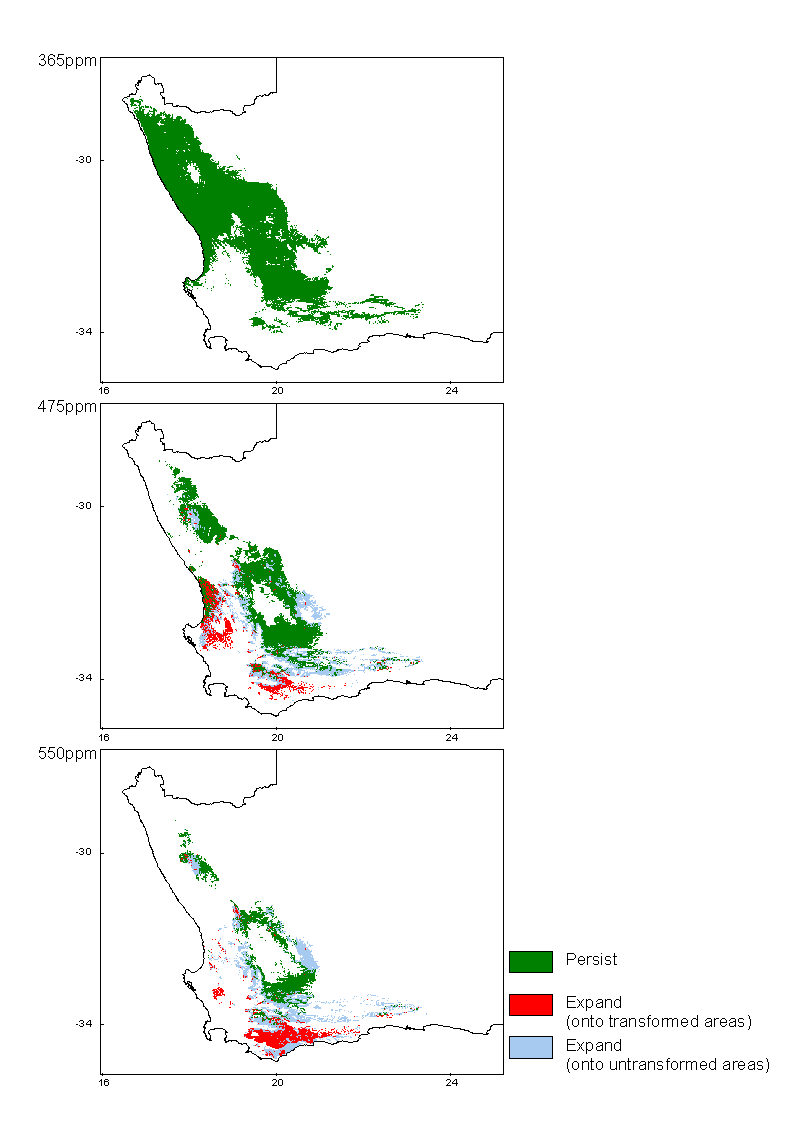
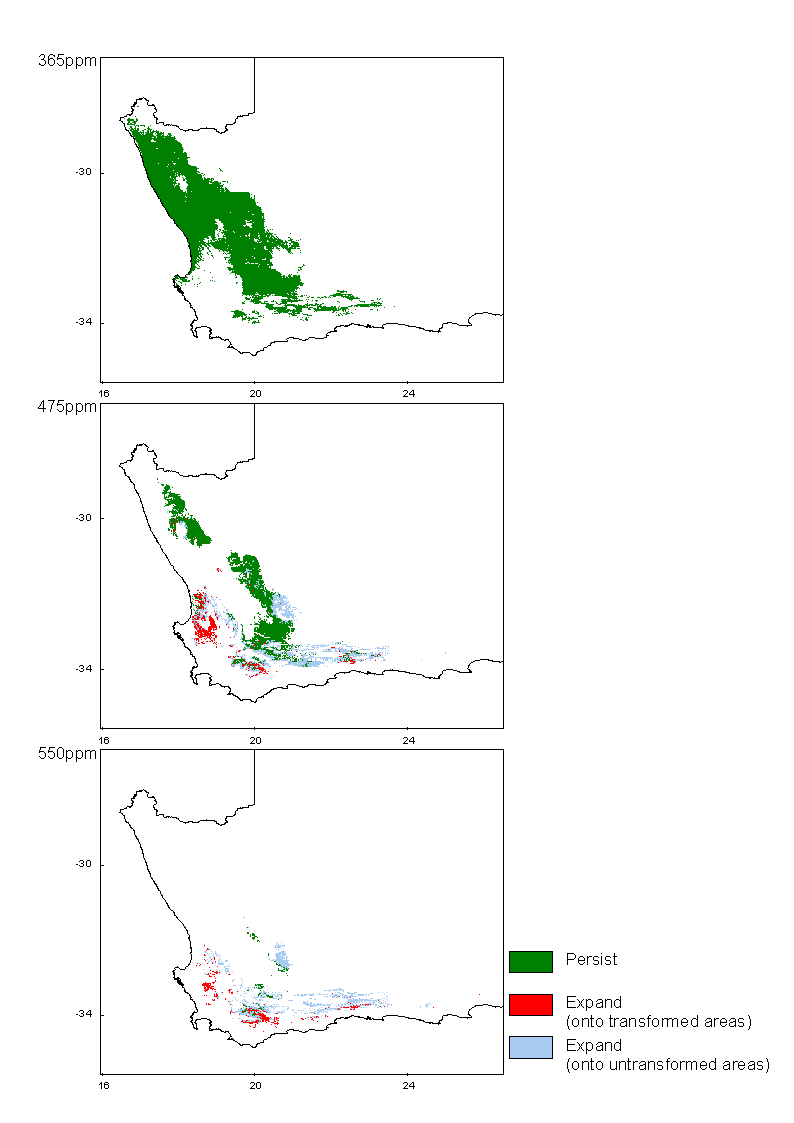
Methods
We used a bioclimatic modeling approach (e.g. Busby 1991) to derive an apparent climate envelope (that is, a "realized niche", [Cao 1995]) for the Succulent Karoo Biome as it exists under current climate conditions. Essentially, bioclimatic modeling determines the environmental limits of an entity with a given spatial distribution by matching its known distribution to climatic surfaces. This is done at a spatial resolution sufficient to avoid ambiguity due to local spatial and topographic heterogeneity.
The spatial distribution of the Succulent Karoo Biome (and the other dominant biomes of South Africa) was digitized from the map presented in Rutherford and Westfall (1994). A climatic database at a spatial resolution of 1 x 1 minute (approximately 1.2 x 1.2 km at this latitude) provided temperature and water availability data for the entire area of interest.
We used as environmental determinants five relatively independent climatically-derived parameters we consider critical to plant physiological function and survival. These are mean minimum temperature of the coldest month (Tmin), heat units (annual sum of daily temperatures (°C) exceeding 18°C [HU18]), Annual Potential Evaporation (PE), Winter soil moisture days (SMDwin), and Summer soil moisture days (SMDsum). Soil moisture days are defined as those days on which soil moisture is above a critical level for plant growth. All of the above parameters were among those derived for South Africa by Schulze and Perks (1999). Climate envelopes were defined simply by the upper and lower limits of each parameter, which is an approximation of the species' realized niche space or bioclimatic envelope (Rutherford et al. 1995, 1996). While there is little experimental work to rely on locally, we consider these five environmental parameters as the minimum basic set for defining a bioclimatic envelope in South Africa's indigenous flora. Tmin is likely to discriminate between species based on their ability to assimilate soil water and nutrients, and continue cell division, differentiation and tissue growth at low temperatures (lower limit), and chilling requirement for processes such as bud break and seed germination (upper limit). Heat units discriminate between species based both on their requirement for a minimum temperature to complete growing cycles (lower limit), and ability to tolerate excess tissue temperature (upper limit). PE discriminates through processes related to transpiration-driven water flow through the plant, and xylem vulnerability to cavitation and water transport efficiency. Soil moisture days in winter and summer discriminate through determining the intensity and duration of seasonal drought stress and growing opportunities, which are especially crucial for seedling recruitment and the survival of this sensitive plant life stage. It should be noted that results are affected by choice of months which define seasonality - in the current study winter was represented by May to August, and summer by November to February.
Predictions of future climate change utilised climate scenarios generated by the widely-used general circulation models CSM and HadCM2, interpolated for South Africa (Schulze and Perks 1999). The HadCM2 model produced two scenarios by using different predictions of warming amelioration by sulphate emissions - these are referred to simply as HadCM2 with, and HadCM2 without sulphates. All models generated predictions of temperature and precipitation change for southern Africa for the mid 21st century (~2050), equivalent to an atmospheric CO2 concentration of 550ppm. Expected rates of change are not given by Schulze and Perks (1999), but in general, more or less linear rates of temperature change are assumed for the next 50 years (Hulme 1996). This assumption allows results for intermediate CO2 levels to be interpolated from the endpoints.
References
Busby JR 1991. A bioclimatic analysis and prediction system. In Nature Conservation: cost effective biological surveys and data analysis, eds, Margules CR, Austin MP. CSIRO Australia pp. 64-68.
Cao G 1995. The definition of the niche by fuzzy set theory. Ecological Modelling 77:65-71.
Hulme M. (ed.) 1996. Climate change and Southern Africa: an exploration of some potential impacts and implications in the SADC region. A Report commissioned by WWF International and co-ordinated by the Climatic Research Unit, UEA, Norwich, UK. Published by the University of East Anglia, Norwich, UK and WWF International, Gland, Switzerland.
Rutherford MC, O'Callaghan M, Hurford JL, Powrie LW, Schulze RE, Kunz RP, Davis GW, Hoffman MT, Mack F 1995. Realized niche spaces and functional types: a framework for prediction of compositional change. Journal of Biogeography 22:523-531.
Rutherford MC, O'Callaghan M, Powrie LW, Hurford JL, Schulze RE 1996. Predicting survival in new environments through analytical GIS application. Environmental Software 11, 113-121.
Rutherford MC, Westfall RH 1994. Biomes of southern Africa - an objective categorization. Second Edition. Memoirs of the Botanical Survey of South Africa 63: 1-94.
Schulze RE, Perks LA 1999. Assessment of the impact of climate: A report to the South African Country Studies Climate Change Programme. School of Bioresources Engineering and Environmental Hydrology. University of Natal, Pietermaritzburg, South Africa.
![]()